|
Dr Manel Baradad. Médico Adjunto. Servicio de Dermatología. Hospital Arnau de Vilanova. Profesor Asociado de Dermatología. Universitat de Lleida
EPIDERMIS
Es un epitelio plano poliestratificado queratinizante que contiene células de diversas estirpes. Tiene un grosor de 0.3 a 1.5 mm. Tridimensionalmente, se proyecta hacia la dermis en forma de crestas engastadas entre las papilas. Su actividad principal es la maduración de queratinocitos (queratinización) desde la capa basal proliferativa, la cual muestra una escasa síntesis de queratina, hasta la capa córnea arregenerativa, donde las células se observan completamente queratinizadas.
Componentes celulares
- Queratinocitos (90-95% de las células): derivados del ectodermo, intervienen en la síntesis de queratina (queratinización) y tienen también funciones inmunes, pudiendo sintetizar citocinas (IL-1, IL-6, IL-8, etc.) en respuesta a diferentes estímulos y presentar antígenos.
- Células de Langerhans (4-5%): derivadas de células madres de la médula ósea (sistema monocito-macrofágico), son procesadoras y presentadoras de antígenos.
- Melanocitos (1%): derivados de la cresta neural o neuroectodermo; son las encargadas de sintetizar melanina (melanogénesis).
- Células de Merkel: forman parte del sistema neuroendocrino; se situan en la capa basal, actúan como mecanoreceptores y están asociadas a terminaciones nerviosas intraepidérmicas.
Estratos de la epidermis
a) Estrato basal o germinativo.
b) Estrato espinoso o de Malpigio.
c) Estrato granuloso.
d) Estrato córneo.
|
|
|

|
| | | | | | |
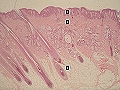
Capas cutáneas. 1: epidermis. 2: dermis. 3: hipodermis
|
Capas cutáneas. 1: epidermis. 2: dermis. 3: hipodermis
|
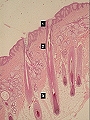
Estratos de la epidermis. 1: basal. 2: espinoso. 3: granuloso. 4: córneo. Papilas dèrmicas (5) y crestas interpapilares (6)
|
Representatión esquemática de los estratos epidérmicos (rojo: melanocito; azul: célula de Langerhans)
| |
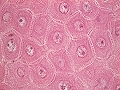
Estrato basal
Llamado también estrato germinativo porque es donde se generan nuevos queratinocitos por proliferación celular, aunque también estan presentes otros tipos de células propias de la epidermis, formando todas ellas una capa única. Los queratinocitos de esta capa se observan como células cuboidales, basófilas, en contacto con y unidas a la membrana basal. Contienen tonofibrillas de citoqueratina, estan fijados en la membrana basal por hemidesmosomas y entre sí por desmosomas y muestran algunos signos de diferenciación (citoqueratinas 5 y 14 y, cuando “ascienden”, 1 y 10). En este estrato, proliferan y desde él ascienden progresivamente hacia los estratos superiores (es decir, hacia la superfície del cuerpo). En conjunto, cerca de un 4% de los queratinocitos basales están en mitosis simultáneamente (la actividad mitótica ocurre principalmente de 00:00 a 04:00h am). Se han caracterizado tres tipos de queratinocitos en esta capa, definidos por el análisis de su cinética: células madre, células amplificadoras transitorias y células comisionadas. Las células madre, que representan un 10% de la población celular basal, tienen una elevada capacidad de división pero son de ciclo largo y débil actividad mitótica; en estudios de proliferación con timidina tritiada, su núcleo la conserva por largos períodos (se las denomina también “células que retienen marcador”). Su división origina las células amplificadoras transitorias; son aproximadamente un 40% de la población queratinocitaria de este estrato y se replican con mucha mayor velocidad que las células madre, aunque son capaces de un número muy limitado de divisiones, de tres a seis, y pierden potencial proliferativo en cada una de ellas. El producto de la última división de las células amplificadoras es la célula comisionada hacia la diferenciación terminal; una célula que ya no prolifera, se diferencia (muestran ya una queratinización incipiente), se desprende de la membrana basal y migra hacia las capas superiores. Finalmente, siendo ya una célula muerta, y pasados unos 60 dias desde la mitosis de una célula madre, se eliminará hacia el entorno con la descamación de la capa córnea.
El carácter indiferenciado de las células madre dificulta la caracterización de marcadores bioquímicos que permitan identificarlas. Se han señalado como posibles marcadores la citoqueratina 19, la mayor expresión de las integrinas beta-1 y alfa-6. Y como probablemente específico el factor de transcripción p63. No asientan únicamente en la capa basal; se han identificado asimismo en el aparato pilosebáceo.
Un defecto en la síntesis correcta de las citoqueratinas de esta capa dará lugar a un defecto congénito de la adhesión entre las células epidérmicas con formación de ampollas y erosiones (p.e., epidermolisis ampollosa simple).
Estrato espinoso o de Malpigio
Situado por encima de la capa basal y formado por 4-10 filas de células cuboides, eosinófilas, que contienen citoqueratinas 1 y 10. Están unidas entre sí por desmosomas, que se observan en los cortes histológicos como “espinas” que rodean a las células (toman este aspecto por haberse retraído en el curso de las manipulaciones técnicas previas). En esta capa, los queratinocitos se diferencian; los tonofilamentos y las tonofibrillas del citoesqueleto incrementan progresivamente su volumen respecto al contenido citoplasmático y, conforme van ascendiendo hasta el estrato granuloso, las células se van aplanando, con su eje mayor paralelo a la superfície corporal. Expresan diversas glucoproteínas de superficie que son moléculas de adhesión de la família de las cadherinas. Y contienen unas vacuolas rodeadas de membrana (queratinosomas o cuerpos de Odland, de morfología discoide) en cuyo interior se depositan precursores de los lípidos epidérmicos (en forma de bicapas laminares de lípidos).
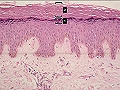
Estrato granuloso
Justo por encima del estrato espinoso se localiza el estrato granuloso. Toma el nombre de su aspecto, que es debido a la presencia de los gránulos de queratohialina, una mezcla de diversas unidades proteicas (junto a las moléculas de queratohialina, la precursora de la queratina, los gránulos contienen prefilagrina). Está formada por 1-2 filas de células aplanadas, basófilas, con abundantes lisosomas que contienen proteasas y nucleasas. En esta capa ocurren la diferenciación final y la muerte celular, con un rápido tránsito a células cornificadas que pierden un 50-85% de su peso, las organelas y el núcleo. Simultáneamente, los cuerpos de Odland liberan polisacáridos, glicoproteínas, hidrolasas ácidas y lípidos al espacio extracelular, elementos que forman, junto a las células muertas, una barrera impermeable a los líquidos.
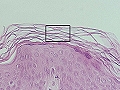
Estrato córneo
Es la capa superior (más externa) de la epidermis, formada por 15-20 capas (más de 100 en palmas y plantas) de células grandes, aplanadas, apiladas como tejas. Son queratinocitos muertos y completamente queratinizados (cornificados, por lo que también se conocen como corneocitos), proteína pura, en definitiva. Constituyen el producto final de la diferenciación epidérmica, cimentados y estabilizados por los lípidos provenientes de los cuerpos de Odland (ceramidas, 40%, ácidos grasos libres, 25%; colesterol, 25%). Entre las células cornificadas se encuentran los lípidos epidérmicos. La queratina es una proteína específica de piel, uñas y pelo. El estrato córneo, especialmente el tercio superior, forma una barrera impermeable que es la verdadera “barrera” cutánea contra los factores exógenos (tóxicos y microorganismos) y la pérdida de agua y electrolitos.
|

|
|
|
| | | | | | |
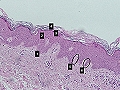
Estrato basal de la epidermis
|
Queratinocitos de la capa basal epidérmica
|
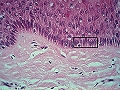
Estrato espinoso de la epidermis
|
Epidermis: 1: estrato granuloso. 2: estrato córneo
|
Estrato córneo de la epidermis
|
Cinética epidérmica
Los queratinocitos muestran una renovación constante que supone la pérdida progresiva de las células del estrato córneo hacia el exterior y su sustitución por células nuevas originadas de la división de células basales, las cuales migran a través de las diversas capas de la epidermis al tiempo que experimentan una diferenciación conocida como queratinización. Durante un período de semanas, las células nuevas pasan desde la capa basal hacia la superfície, mostrando cambios en su forma, desde la columnar original en la base hacia cuboidal y finalmente aplanada; simultáneamente, se modifica la composición y agregación de las citoqueratinas intracelulares. El recambio completo desde la capa basal hasta la eliminación al exterior de los corneocitos ocurre en unos dos meses (1 mes en capas viables + 1mes en estrato corneo).
La renovación epidérmica es estimulada por diversos factores de crecimiento, entre ellos el factor de crecimiento epidérmico (“EGF”), el factor transformador del crecimiento alfa (“TGF-a”), interleucinas (IL-1, IL-2, IL-6) y otras citocinas, así como por el factor básico de crecimiento de los fibroblastos (“bFGF”). Y es inhibida por por el factor transformador del crecimiento beta (“TGF-b”), los interferones alfa y gamma, y el factor de necrosis tumoral (“TNF”), entre otros.
Citoqueratinas y queratinización
El citoesqueleto es un complejo sistema de proteínas intracitoplasmáticas formado por microfilamentos (6 nm), microtúbulos (25 nm) y filamentos intermedios. Estos últimos constituyen la mayor parte del citoesqueleto y cumplen una variedad de funciones relacionadas con la forma celular, su organizacion espacial y la transferencia de información; también actúan de soporte a mitocondrias, poliribosomas, enzimas, ácidos nucleicos y otros componentes.
Según sus propiedades bioquímicas, biofísicas y antigénicas, se han reconocido diferentes tipos de filamentos intermedios en cada tipo celular: vimentina (células mesenquimales), desmina (células musculares), proteína acídica fibrilar de la glia (astrocitos) y citoqueratinas en las células de estirpe epitelial. En cultivos de células epidérmicas, las citoqueratinas suponen el 30% de las proteínas celulares, mientras que en el estrato córneo suponen el 85%. Son proteinas fibrosas que forman una molécula alfa-helicoidal con tres filamentos de siete polipéptidos que se agregan progresivamente para formar unidades de mayor tamaño (tonofilamentos y tonofibrillas).
Se han identificado 19 tipos de citoqueratinas, con un peso molecular de 40-68 kd y un pH de 5-8, distinguiéndose entre las dos subfamílias: acídica y básica. El peso molecular de la subfamília básica es relativamente mayor que el de los miembros de la acídica. Cada citoqueratina es el producto de un único gen, y en condiciones normales se expresan a pares conteniendo un miembro de cada subfamília. Los dos miembros del par ocupan un lugar similar de sus respectivas famílias ordenadas por tamaño.
El tipo de citoqueratina difiere entre los diferentes tejidos y tambien entre las células de un mismo tejido (pe, es diferente entre las células de proliferantes de la capa basal epidérmica y las células cornificadas del estrato córneo).
En la epidermis, la síntesis de citoqueratinas se inicia en la capa basal (tonofilamentos de citoqueratina 5 y 14), es máxima en el estrato espinoso (tonofibrillas de citoqueratina 1 y 10) y finaliza en el estrato granuloso (filamentos de queratina unidos por filagrina y loricrina). En este estrato tiene lugar la cornificación de los queratinocitos; las células mueren, pierden las organelas citoplasmáticas, el núcleo y el 70% del agua. Se vierten los lípidos de los cuerpos de Odland al espacio extracelular, la membrana plasmática se refuerza con involucrina, el citoesqueleto se condensa en una masa amorfa de queratina, filagrina y loricrina. El resultado final es un queratinocito que ha alcanzado su destino, la completa diferenciación en corneocitos, en escamas córneas.
La queratinización o la cornificación incorrectas dan lugar a diversas dermatosis (queratodermias palmoplantares, ictiosis, etc).
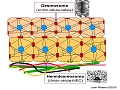
Desmosomas
Son estructuras especializadas en la unión intercelular. Con el microscopio electrónico se observan como placas electrón-densas en la membrana de los queratinocitos. En su porción intracelular se identifican desmoplaquina I y II (molécula de adhesión de la família de las cadherinas), envoplaquina, periplaquina y placofilina; en conjunto anclan la estructura desmosómica al citoesqueleto. Su porción transmembrana contiene desmogleína y descomolina, que se unen a los péptidos homólogos de las células vecinas.
En algunas enfermedades, los desmosomas no se forman bien o se alteran por la acción de autoanticuerpos, lo que se traduce en una pérdida de cohesión de los queratinocitos (acantolisis) y la aparición de ampollas que se rompen fácilmente (pénfigos).

|

|
|
|
| | | | | | |
Representación esquemática de un desmosoma
|
Estructura del desmosoma
|
Estructuras de unión de la piel
| | |
Células dendríticas epidérmicas
Melanocitos
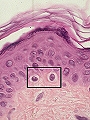
Son células mesenquimales que derivan de la cresta neural. En el embrión migran precozmente desde aquella hacia la epidermis y los folículos pilosebáceos. Se sitúan en la capa basal epidérmica, en contacto con los queratinocitos, a razón de uno por cada 30-40 queratinocitos, formando en conjunto la “unidad melanoepidérmica”. Su número varía según la región anatómica, pero es constante entre las personas de las diferentes razas; contrariamente a la creencia popular, las diferencias en la pigmentación racial no son debidas al número de melanocitos sino al número y tamaño de los melanosomas que producen.
Tienen una morfologia cuboide, poseen dendritas que se extienden entre los queratinocitos, carecen de desmosomas y se observan de color claro en las preparaciones histológicas teñidas con h&e (figura 10). Pueden detectarse con la tinción de Fontana-Masson, la reacción de la Dopa de Bloch y mediante anticuerpos monoclonales (S-100 y HMB-45 +).
Sintetizan el pigmento (melanina) en organelas especiales dotadas de membrana (melanosomas). Se parte de la fenilalanina =/1/=> tirosina =/2/=> DOPA =/3/=> DOPAquinona =/4/=> melanina ==> agregados de melanina/maduración de los melanosomas. La enzima encargada de los pasos 2 y 3 es la tirosinasa, cuya transcripción y síntesis están reguladas por la acción de la hormona melanocitoestimulante (“MHS”) sobre su receptor de membrana y a través de la vía de señales transmembrana de la proteín-quinasa. Existen dos tipos de melanina, eumelanina (de color marrón pardo) y feomelanina (rojo amarillo). Se produce un tipo u otro según la variante (existen 5) de receptor para MSH presente en la superficie celular.
Terminada la maduración de los melanosomas, se desplazan hacia los extremos de las dendritas, desde donde son capturados por los queratinocitos mediante un proceso de fagocitosis. En los queratinocitos situados más interiormente, los gránulos de melanina se distribuyen formando un “sombrero” protector sobre la parte externa del núcleo. En la capa córnea, están distribuidos uniformemente formando un “manto” que reduce la cantidad de radiación que penetra en la piel.
La radiación ultravioleta, especialmente el espectro B (290-320 nm.), oscurece la piel inicialmente por fotooxidación inmediata de la melanina ya formada y estimulando la síntesis de nueva melanina, que será evidente tras un periodo de días. Asimismo, la radiación UV provoca un engrosamiento de la epidermis induciendo proliferación de los queratinocitos, que también se traduce como oscurecimiento cutáneo.
La melanina protege de la radiación ultravioleta y por tanto de sus efectos agudos y crónicos. Su ausencia por defecto en la síntesis (albinismos) o pérdida de melanocitos (vitíligo) origina una mayor susceptibilidad a las quemaduras solares y facilita la carcinogénesis.
Célula de Langerhans
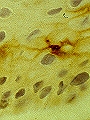
Se originan en células madre de la médula ósea y pertenecen al sistema monocito-macrofágico. Se sitúan distribuidas homogéneamente en las capas suprabasales y el estrato espinoso. Su citoplasma es claro y poseen ramificaciones dendríticas. Observadas al microscopio electrónico, poseen unas organelas específicas, los gránulos de Birbeck (en forma de raqueta). Son ATPasa+ y se pueden identificar tambien por la detección de diferentes antígenos (CD1, CD4, S100, HLA-DR, receptor de C3 y de Fc de la IgG).
Tienen una función inmunológica: captación, procesamiento y presentación de antígenos a los linfocitos T para iniciar la respuesta inmune. También sintetizan citocinas (IL-1, IL-12, FNT-alfa,...)
En conjunto, forman la primera línea de defensa inmunológica frente a los antígenos ambientales (pe, inician la reacción que conduce a las dematitis alérgicas de contacto). Y son células capitales en la inmunovigilancia frente a infecciones y carcinomas cutáneos.
Célula de Merkel
Se cree son células epiteliales neuroendocrinas. Están situadas entre los queratinocitos de la capa basal; tienen un citoplasma claro con gránulos que contienen neuropéptidos, filamentos de citoqueratina (es muy específica de ellas la 20) y poseen un núcleo lobulado. Proyectan dendritas y forman desmosomas. Están asociadas a terminaciones nerviosas intraepidérmicas. Se localizan en las regiones con mayor sensibilidad táctil como los labios, pulpejos de dedos y folículo pilosebáceo.
Son mecanoreceptores de adaptación lenta que reciben información sobre la deformación de las células epidérmicas.
|
|
|
|
| | | | | | |
Melanocito: célula clara entre los queratinocitos basales
|
Célula de Langerhans en el estrato espinoso, identificada mediante ac monoclonales (S-100)
| | | |
|