|
Dra Roser Tamarit. Servei de Dermatologia. Hospital de la Ribera. Valencia. Dr Joan Ribera. Catedrático de Biología. Universitat de Lleida.
INTRODUCCIóN: MECANISMOS DE ADHESIóN DE LA PIEL
El cuerpo humano es el resultado de una agrupación ordenada de células perfectamente ensambladas que, por lo general, mantienen una localización precisa. Las distintas agrupaciones de células conforman los tejidos de nuestro organismo y su ensamblaje se produce mediante moléculas y estructuras, ubicadas en la membrana, responsables de la adhesión celular. La adhesión celular es asimismo el mecanismo que emplean las células para realizar los procesos migratorios que se producen durante el desarrollo o la reparación de los tejidos.
Moléculas de adhesión celular (MAC)
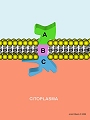
Son proteínas transmembranarias ubicadas en la membrana celular que tienen tres dominios: un dominio extracelular a través del cual interaccionan con la superficie de otras células o componentes de la matriz extracelular, un dominio hidrofóbico, situado en la región central de la molécula, a través del cual se anclan a la membrana, y un dominio intracelular, en contacto con la región citoplasmática, a través del cual interacciona con diferentes proteínas que asocian las MAC al citoesqueleto de la célula, estableciéndose una continuidad morfofuncional entre el citoesqueleto y la superficie de las células.
Las MAC son las moléculas responsable de la adhesión de una célula a las de su entorno y a la matriz extracelular. Además participan en procesos de comunicación intercelular. El intercambio de información entre células cercanas se consigue gracias a la acción de citocinas sobre receptores específicos. Para conseguir dicho intercambio se precisa que las células se mantengan unidas, para lo cual intervienen las moléculas de adhesión. Su papel es asimismo fundamental en reacciones inmunes y también en la organización estructural de los tejidos durante la embriogénesis.
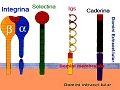
Existen 4 grandes familias de MAC: las caderinas, las selectinas, las integrinas y la superfamilia de las inmunoglobulinas. Las integrinas y las caderinas son las que tienen un papel más relevante en la organización de la piel.
Las caderinas son las primeras moléculas de adhesión que aparecen durante el desarrollo embrionario. Tienen una gran importancia en la compactación, cambio morfológico que se produce a partir de que el embrión tenga 8 células. El cambio consiste en la agrupación celular mediante interacciones establecidas entre las E-caderinas presentes en la superficie celular. Las caderinas son proteínas diméricas que se caracterizan por tener un dominio extracelular muy voluminoso formado por 5 segmentos repetidos. Estos dominios para ser funcionales se han de unir a calcio y a través de ellos la célula interacciona con las caderinas del mismo tipo de las células vecinas. Este tipo de unión, en el que la misma molécula actúa como receptor y ligando se denomina homofílica. Por su dominio intracelular habitualmente interaccionan con elementos del citoesqueleto mediante una serie de proteínas puente, estableciendo así una continuidad morfofuncional entre las células que se mantienen unidas.
En los queratinocitos se encuentran dos tipos de caderinas, las desmocolinas y las desmogleinas; que están especialmente concentradas en los desmosomas.
Las integrinas son proteínas transmembranarias heterodiméricas que rolex nachbau están formadas por dos cadenas polipeptídicas, α y β. Para que puedan ser funcionales han de estar unidas a cationes divalentes (Ca2+ o Mg2+). Las integrinas humanas resultan de la combinación de 9 cadenas de tipo β con 24 de tipo α y por su dominio extracelular interaccionan, con componentes de la matriz extracelular (MEC) mientras que por su dominio intracelular lo hacen con elementos del citoesqueleto (microfilamentos o filamentos intermedios) gracias a una serie de proteínas puente. Con ello se establece una continuidad morfofuncional entre el citoesqueleto de la célula y la MEC. En los queratinocitos, la integrina α6β4 constituye el principal componente de los hemidesmosomas, estructura a través de la cual los queratinocitos basales se adhieren a la lamina basal de la matriz extracelular. Las integrinas, además, activan vías de transducción de señales intracelulares.
|
|
|
|
| | | | | | |
Estructura transmembrana de una molécula de adhesión
|
Moléculas de adhesión epiteliales
|
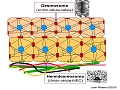
Estructuras de unión de la piel
| | |
Estructuras celulares responsables de la adhesión celular intraepidèrmica: desmosomas
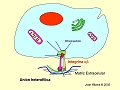
Los queratinocitos se mantienen unidos entre sí gracias a los desmosomas, estructuras especializadas en la unión intercelular. Si observamos la epidermis al microscopio óptico,replique montre los desmosomas se presentan como finos puentes que conectan los queratinocitos suprabasales entre sí, lo que les proporciona un aspecto “espinoso”. Al microscopio electrónico se ven como regiones diferenciadas de la membrana en zonas donde las células están muy próximas (a menos de 100 nm). Constan de unas estructuras electrodensas en forma de disco (placa densa) en su cara citoplasmática, y de unas proteínas transmembranarias denominadas caderinas que, desde la placa densa, parten hacia el exterior de la célula y dependen de calcio para ser funcionales. Entre las caderinas epidérmicas cabe destacar las desmogleinas (I y III), las desmocolinas (I-IV) y una proteina de 130 kD emparentada con la desmogleina 1, que forman agregados macromoleculares en el exterior de la célula. Por su porción citoplasmática interaccionan con la placoglobina, las placofilinas y la desmoplaquina, proteínas que actúan de puente entre las caderinas y el citoesqueleto celular (filamentos de queratina) y forman la placa densa.
Los desmosomas también intervienen en la morfogénesis de la epidermis y en la transmisión de señales desde la membrana plasmática y el citoesqueleto hacia el núcleo. También se ha descrito la existencia de caderinas en otros tejidos como el neural y el placentario.
La alteración de alguno de los componentes que forma parte del desmosoma, ya sea por autoanticuerpos como en los pénfigos, o bien por alteraciones estructurales genéticas o adquiridas, da lugar a la formación de dehiscencias que originan ampollas intraepidérmicas con células flotando en su interior, fenómeno al que se conoce como acantolisis. La unión de los anticuerpos a las desmogleinas y desmocolinas debilita las interacciones entre estas caderinas y en consecuencia las células se separaran con mucha facilidad.

|

|
|
|
| | | | | | |
Representación esquemática de un desmosoma
|
Estructura del desmosoma
|
Estrato espinoso de la epidermis. Las "espinas" corresponden a los desmosomas
|
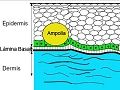
Formación de una ampolla intraepidérmica
| |
Mecanismos de unión dermoepidérmicos: región de la membrana basal, hemidesmosomas y fibrillas de anclaje
Ultraestructuralmente, la región de la membrana basal consta de 2 compartimientos. El primero está compuesto por la membrana plasmática de los queratinocitos de la capa basal epidérmica, en su parte superior, la lámina densa por debajo y el espacio claro que queda entre ambos que se conoce como lámina lúcida. El segundo es el situado por debajo de la lámina densa, denominado lámina subbasal. La lamina densa (membrana basal) es una región especializada de la matriz extracelular sobre la cual descansan los epitelios. Está constituida fundamentalmente por colágeno IV, que polimeriza formando mallas tridimensionales sobre las cuales se depositan los proteoglicanos y otras proteínas como la laminina. El papel mas relevante de la lámina basal es la de intervenir en la compartimentación de los tejidos que forman un órgano, estableciendo una clara separación entre las células parenquimatosas y el tejido conectivo. En órganos como en el pulmón constituye una separación entre poblaciones celulares distintas. Por lo general la lámina basal actúa como una barrera que impide la migración de células entre los diferentes compartimentos que delimita.
En el primer compartimiento se localizan los hemidesmosomas, estructura de adhesión celular que une los queratinocitos de la capa basal de la epidermis a una región especializada de la matriz extracelular, la lámina basal. Con el microscopio electrónico se ven como la mitad de un desmosoma, de lo que deriva su nombre. Presentan una placa electrodensa, en la cara citoplasmática de la célula, sobre la cual se anclan filamentos de queratina. En esta región se encuentra concentrada la integrina α6ß4, molécula de adhesión celular que interacciona por su región extracelular con la laminina 5, componente de la lámina basal sobre el que descansa la epidermis. Como en los desmosomas de las capas superiores de la epidermis, la integrina α6β4 por su dominio intracitoplasmático interacciona con un conjunto de proteínas que forman un agregado macromolecular que se conoce como placa densa. De estas proteínas la más conocida es la plectina, que actúa como elemento agregante de otras proteínas alrededor de las integrinas. El agregado macromolecular resultante interacciona con los filamentos de queratina del citoesqueleto dando consistencia y estabilidad al complejo. Otras glucoproteínas transmembranarias que forman parte de los hemidesmosomas son las BP 230 y 180, conocidas así por ser los antígenos del penfigoide I y II de 230 y 180 kD respectivamente. Están compuestos de colágeno XVII y pertenecen a las caderinas de unión célula-matriz.
En el segundo compartimiento, la lámina subbasal, se encuentran las fibrillas de anclaje, auténticas responsables de la unión de la epidermis a la dermis, compuestas de colágeno tipo VII. Dichas fibrillas se unen en su parte superior con la membrana basal y en la inferior se entrelazan con las fibras de colágeno I y III de la dermis. También se entrelazan entre sí formando placas de anclaje, constituidas por colágeno IV.
Existe un grupo de enfermedades autoinmunes que conocemos como Penfigoides, en que las personas afectadas sintetizan autoanticuerpos contra las proteínas BP180 y BP230 del hemidesmosoma. Esta reacción debilita la unión de las células a la matriz extracelular y convierten la interfase célula-matriz extracelular en una región muy frágil que se rompe con facilidad, separándose la dermis de la epidermis lo que origina una ampolla. Por otra parte en el lupus eritematoso ampolloso y en la epidermolisis ampollosa adquirida se sintetizan autoanticuerpos anticolágeno tipo VII, lo que 3621px;">rolex replica aunque en este caso se localizará subbasal. También se producen ampollas subepidérmicas en caso de déficits estructurales congénitos de proteínas de unión de esta región. En la epidermolisis ampollosa distrófica se ha detectado una reducción del colágeno tipo VII, por mutaciones del gen del mismo. Desde el nacimiento los niños que padecen esta mutación tienen una gran fragilidad de la piel, lo que origina ampollas y erosiones en las zonas de mínimos traumatismos, que se mantiene de por vida.

|
|

|

|
| | | | | | |
Región de la membrana basal, esquema (en azul, elementos de la membrana basal)
|
Representación esquemática de un hemidesmosoma
|
Estructura del hemidesmosoma
|
Formación de una ampolla subepidérmica
| |
|